Do, 25.10.2012- 04:20 — Walter Jakob Gehring
![]()

"Die Wissenschaft übertrifft Hollywood“ titelte die New York Times im März 1995, als Walter Gehring mit dem Gen Pax6 den Hauptschalter in der Entwicklung des Auges entdeckt hatte und dessen Funktion durch die Generierung zusätzlicher Augen auf unterschiedlichen Körperteilen der Taufliege bewies. Spätere Untersuchungen Gehrings ergaben, daß das Pax6-gesteuerte Augenentwicklungsprogramm ein universelles, bei allen Tieren von den Plattwürmern bis zu den Säugetieren, vorliegendes Prinzip darstellt, der Ursprung all der unterschiedlichen Augentypen also vom selben Prototyp ausgeht.
Die Natur hat im Verlauf der Evolution die verschiedensten Augentypen hervorgebracht. Diese reichen vom Kameraauge der Wirbeltiere mit einer Linse, die das einfallende Licht auf eine lichtempfindliche Netzhaut (Retina) projiziert, über das komplexe Facettenauge der Insekten und anderer Gliederfüßler, das aus zahlreichen Einzelaugen mit je einer Linse und einer Gruppe von Photorezeptorzellen zusammengesetzt ist, bis zum Spiegelauge, das beispielsweise bei der Jakobsmuschel sowohl über eine Linse als auch über einen reflektierenden Parabolspiegel verfügt, die das Licht auf eine Netzhaut projizieren. Abbildung 1.
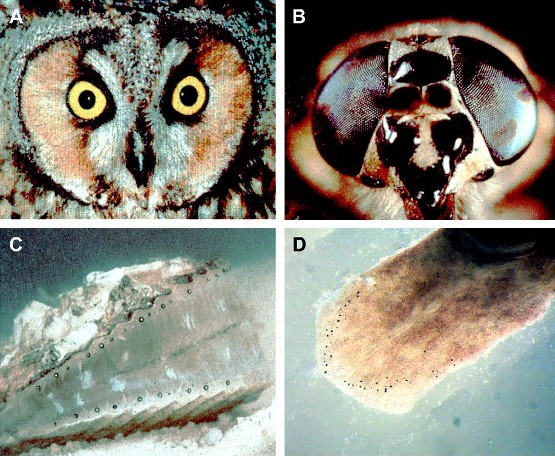 Abbildung 1. Unterschiedliche Augentypen. A) Kamera-Auge der Eule, B) Facettenauge der Fliege, C) Spiegel-Auge der Jakobsmuschel, D) Prototyp-Auge bestehend aus einer Pigmentzelle und einer Photorezeptorzelle des Plattwurms Polycelis auricularis.
Abbildung 1. Unterschiedliche Augentypen. A) Kamera-Auge der Eule, B) Facettenauge der Fliege, C) Spiegel-Auge der Jakobsmuschel, D) Prototyp-Auge bestehend aus einer Pigmentzelle und einer Photorezeptorzelle des Plattwurms Polycelis auricularis.
Trotz dieser morphologischen Vielfalt bestimmen ähnliche physikalische und chemische Prinzipien den Sehvorgang. In allen Vielzeller-Organismen finden sich sogenannte Opsine als Sehpigmente. Dies sind Photorezeptor-Proteine, die - von einem einzelnen Lichtimpuls (Photon) angeregt - eine Konformationsänderung durchlaufen und damit einen Nervenimpuls auslösen.
Darwins Hypothese zum Ursprung des Auges
Für Darwin war es eine besondere Herausforderung, die Entstehung und Entwicklung des Auges zu erklären, und er widmete diesem Thema ein ganzes Kapitel in seinem Buch „The Origin of Species“. Er war davon überzeugt, daß ein so perfektes Organ wie beispielsweise das Auge eines Adlers nicht ein durch rein zufällige Variationen und Selektion entstandener Prototyp sein könnte, weil ja Selektion erst einsetzen kann, wenn ein bereits ansatzweise funktionierender Prototyp existiert. In genialer Weise fand er eine Lösung des Problems: Er postulierte, daß sehr früh in der Entwicklung ein ganz einfaches, aus zwei Zellen - einer Nervenzelle (Photorezeptorzelle) und einer Farbstoffzelle (Pigmentzelle) - bestehendes Auge existiert haben mußte, das ausreichte um seinem Träger bereits das Richtungssehen und damit einen Selektionsvorteil zu ermöglichen. Ausgehend von diesem höchst einfachen Prototyp hätten sich dann die viel komplexeren Augen eines Insekts, eines Fisches, eines Vogels entwickelt.
Tatsächlich konnte später ein derartiger Prototyp bei bestimmten Plattwürmern in Japan gefunden werden (Abbildung 1 D). Derartige Neuentwicklungen sind in der Evolution sehr seltene, zufallsbedingte Ereignisse.
Sind die verschiedenen Augentypen aus einem Prototyp oder voneinander unabhängig entstanden?
Neodarwinisten unterlegten Darwins Theorie der „Entstehung der Arten“ mit genetischen Prinzipien. Da die verschiedenen Augentypen einen unterschiedlichen morphologischen Aufbau haben und sich embryologisch unterschiedlich entwickeln, wurde bis in die jüngste Zeit angenommen, daß die Augen in den 40 bis 60 verschiedenen Tiergruppen voneinander unabhängig – polyphyletisch - entstanden seien. Das klassische Dogma, das man zur Zeit noch in fast allen Lehrbüchern findet, besagt, daß das Komplexauge der Insekten und das Linsenauge der Wirbeltiere keine gemeinsamen Vorfahren hätten.
Da sich die Entstehung derartiger Prototypen aber nicht durch Selektion erklären läßt, müßten diese sehr seltenen und zufälligen Ereignisse 40 bis 60 Mal unabhängig voneinander eingetreten sein.. Unsere neueren molekulargenetischen Untersuchungen haben jedoch gezeigt, daß die verschiedenen Augentypen, die man im Tierreich findet, alle durch das gleiche Masterkontrollgen – eine Art Hauptschalter – namens Pax6 gesteuert werden und somit – monophyletisch - auf einen gemeinsamen Ursprung zurückgehen.
Pax6 – essentiell für die Entwicklung des Auges
Welche Gene einen Einfluß auf die Entwicklung des Auges haben, läßt sich auf Grund auftretender Mutationen nachweisen: Bei der Maus wurde eine Mutation („small eye“) beschrieben, die bei mischerbigen Tieren (mit einer defekten und einer funktionstüchtigen Kopie des Gens) zu einer Reduktion der Augen führt. Bei reinerbigen Embryonen (mit zwei defekten „small eye“-Genen von beiden Eltern) werden keine Augen gebildet, aber auch keine Nase und ein großer Teil des Gehirns fehlt – die Embryonen sterben bereits im Uterus. Ein ähnlicher, als Aniridia (= fehlende Iris) bezeichneter Erbdefekt findet sich auch beim Menschen: Individuen mit nur einem funktionstüchtigen Gen bilden im Extremfall keine Iris, sind also blind. Zwei abortive – vermutlich reinerbige Föten hatten, wie bei der Maus keine Augen, Nasen und reduzierte Gehirne.
Die mutantentragenden Gene wurden zuerst bei der Maus und anschließend beim Menschen kloniert und als sogenanntes Pax6-Gen identifiziert; ein bei der Taufliege Drosophila gefundenes homologes Pax6-Gen entsprach der bereits lange als „eyeless“ beschriebenen Mutante. Abbildung 2 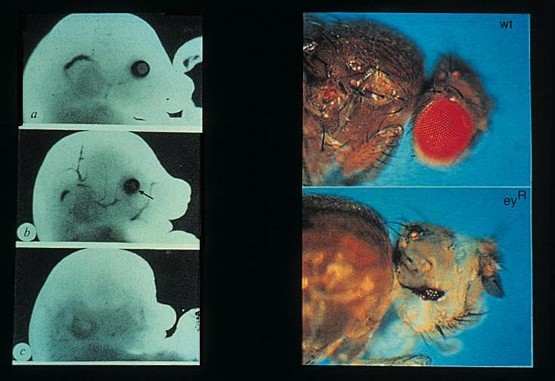 Abbildung 2. Die Mutationen des Pax6-Gens. Links: Small-eye bei der Maus (a: normaler Mausembryo, b: mischerbiger Small-eye-Typ, c: reinerbiger Small-eye Typ), rechts eyeless bei der Taufliege (oben: normaler Kopf, unten reinerbiger eyeless-Typ).
Abbildung 2. Die Mutationen des Pax6-Gens. Links: Small-eye bei der Maus (a: normaler Mausembryo, b: mischerbiger Small-eye-Typ, c: reinerbiger Small-eye Typ), rechts eyeless bei der Taufliege (oben: normaler Kopf, unten reinerbiger eyeless-Typ).
Pax6 – Hauptschalter in der Entwicklung und Evolution des Auges
Das Pax6-Gen kodiert für ein Protein, das als Regulator (Transkriptionsfaktor) die Aktivität der ihm untergeordneten Gene steuert und in der Evolution hoch konserviert ist. In der Maus ist dieses Pax6-Protein bereits in den frühesten Stadien der Augenentwicklung – wenn sich das Augenbläschen aus dem Gehirn stülpt – in allen Teilen der Augenanlage exprimiert, ebenso trifft dies für die eyeless-Mutante der Fliege zu.
Diese Übereinstimmung zwischen Wirbeltieren und Wirbellosen brachte mich auf die Idee, es könnte sich bei Pax6 um ein Masterkontrollgen – einen Hauptschalter – in der Augenentwicklung handeln. Den Nachweis ob dieses einzelne Gen fähig ist, die gesamte Kaskade der für die Augenentwicklung benötigten, mehr als tausend Gene zu induzieren, versuchten wir durch gezielte „ektopische“ Expression von Pax6 zu erbringen, d.h. durch Expression nicht nur in den Augen sondern beispielsweise in Beinen, Flügeln, Antennen der Taufliege. Da der Entwicklungsgang dieser Körperteile bereits im frühen dritten Larvenstadium festgelegt wird, mußte Pax6 noch vor diesem Zeitpunkt „angeschaltet“ werden.
Zur großen Überraschung der Fachwelt war dieser Versuch erfolgreich, es gelang zusätzliche Augen auf den erwähnten Körperteilen zu erzeugen (Abbildung 3). Wie später gezeigt wurde, waren diese Augen auch zum Sehen befähigt. 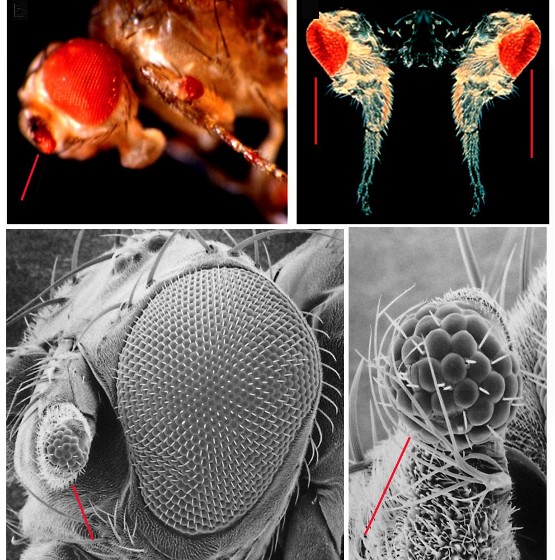 Abbildung 3. Ektopische Induktion zusätzlicher Augen in der Taufliege durch Pax6. Gezielte Expression des mutierten Pax6 Gens „eyeless“ (Drosophila) führt zur Induktion zusätzlicher Augen in den Antennen (oben links) und in den Beinen (oben rechts). Drosophila. Das Maus Pax6-Gen induziert ebenfalls zusätzliche Augen in den Antennen (umten links, rechts in Vergrößerung). Die zusätzlichen Augen sind durch rote Linien markiert.
Abbildung 3. Ektopische Induktion zusätzlicher Augen in der Taufliege durch Pax6. Gezielte Expression des mutierten Pax6 Gens „eyeless“ (Drosophila) führt zur Induktion zusätzlicher Augen in den Antennen (oben links) und in den Beinen (oben rechts). Drosophila. Das Maus Pax6-Gen induziert ebenfalls zusätzliche Augen in den Antennen (umten links, rechts in Vergrößerung). Die zusätzlichen Augen sind durch rote Linien markiert.
Außerdem konnte gezeigt werden, daß auch die Expression des Pax6-Gens der Maus geeignet ist, zusätzliche Augen in ektopischen Stellen der Taufliege zu erzeugen. Die Pax6-Gene von Wirbeltieren und Wirbellosen sind also in ihrer Funktion äquivalent! Diese erstaunlich hohe evolutionäre Konservierung deutet auf einen starken Selektionsdruck und auf die wichtige funktionelle Bedeutung des Gens hin. Zusätzliche Augen wurden auch in Frosch-Embryonen erzeugt, denen das Pax6-Genprodukt der Taufliege injiziert worden war. Schlußendlich wurde das Pax6-Gen in einer Vielzahl an Spezies bis hin zu den Plattwürmern gefunden.
Schlußfolgerungen
Unsere molekulargenetische Untersuchungen haben ergeben, daß das Augenentwicklungsprogramm bei allen Tieren von den Plattwürmern bis zu den Säugetieren, den Menschen miteingeschlossen, übereinstimmt. Rund 65 % der Gene, die in der Retina der Taufliege exprimiert werden, sind auch in der Retina der Maus aktiv. Zuoberst in der Hierarchie steht das Masterkontrollgen Pax6. Bereits der zweizellige Prototyp des Auges, der noch in den Plattwürmern gefunden wird, wurde von Pax6 gesteuert. Diese Steuerung wurde auch bei allen höheren Lebewesen beibehalten, hat aber zu den verschiedensten Augentypen geführt, wobei immer mehr Gene (beispielsweise für die Entwicklung der Linsen) in den Entwicklungsweg eintraten.
Der Ursprung all dieser Typen geht also vom selben Prototyp aus. Die Hypothese Darwins ist damit glänzend bestätigt worden. Abbildung 4.
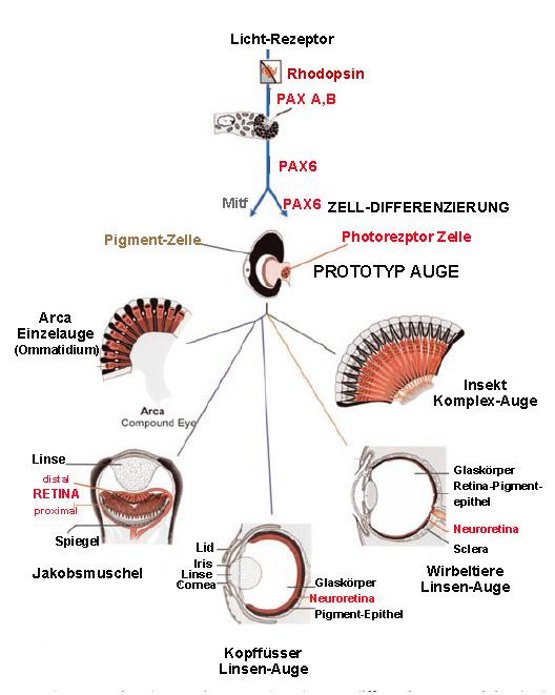 Abbildung 4. Schema der Evolution des Auges. Der erste Schritt ist die Evolution eines Lichtrezeptormoleküls, das bei allen vielzelligen Organismen Rhodopsin ist. Aus der ersten einzelligen Photorezeptorzelle entstand dann durch Pax6- und MITF-gesteuerte Zelldifferenzierung der zweizellige, von Darwin postulierte Prototyp des Auges (MITF ist ein Regulator für die Pigmentzelle). Aus diesem Prototyp haben sich alle komplexeren Augentypen entwickelt, wobei immer mehr Gene(beispielsweise für die Entwicklung der Linsen) in den Entwicklungsweg eintraten.
Abbildung 4. Schema der Evolution des Auges. Der erste Schritt ist die Evolution eines Lichtrezeptormoleküls, das bei allen vielzelligen Organismen Rhodopsin ist. Aus der ersten einzelligen Photorezeptorzelle entstand dann durch Pax6- und MITF-gesteuerte Zelldifferenzierung der zweizellige, von Darwin postulierte Prototyp des Auges (MITF ist ein Regulator für die Pigmentzelle). Aus diesem Prototyp haben sich alle komplexeren Augentypen entwickelt, wobei immer mehr Gene(beispielsweise für die Entwicklung der Linsen) in den Entwicklungsweg eintraten.
Weiterführende Links
PBS - Genetic Toolkit 4:47 min (Englisch)
International workshop on Evolution in the Time of Genomics (7 – 9 May 2012)- part 07. 1:27:37. Vortrag von Walter Gehring rund 30 min (ab: 38 min). (in Englisch; setzt etwas an biologischen Grundkenntnissen voraus)
- Printer-friendly version
- Log in to post comments