Fr, 05.01.2012 - 04:20 — Inge Schuster

![]()
Dieser Beitrag dient als Einleitung zum Artikel von Wolfgang Knoll: „Die biomimetische künstliche Nase – wie weit sind wir?“, der in mehreren Teilen in den nächsten Wochen erscheinen wird.
Der letzte Beitrag von Gottfried Schatz im Science-Blog „Wie Gene und chemische Botenstoffe unser Verhalten mitbestimmen“ hat sich mit der Kommunikation von Nervenzellen beschäftigt. Diese erfolgt mit Hilfe kleiner chemischer Moleküle – Botenstoffen – die von einer elektrisch angeregten Senderzelle ausgestoßen werden, an spezifische Rezeptoren von Empfängerzellen andocken und mittels dieser Rezeptoren die Auslösung elektrischer Signale bewirken. Der Signaltransfer hängt von Art, Eigenschaften und individuellen Varianten der Rezeptoren ab, ebenso wie von denen der Proteine, die Synthese und Metabolismus der Botenstoffe bewirken.
Rezeptoren finden sich nicht nur auf Nervenzellen. Alle Zellen, von Einzellern bis zu den Zellen hochkomplexer Organismen, sind auf ihrer Oberfläche mit einer Vielfalt derartiger, hochspezifischer Sensoren ausgestattet, mit deren Hilfe sie miteinander kommunizieren und ebenso. unterschiedlichste Informationen aus ihrer Umgebung aufnehmen und auf deren Reize reagieren. Rezeptoren kontrollieren praktisch alle unsere physiologischen Funktionen, beginnend bei unseren Sinnesempfindungen, indem sie optische und akustische Reize, Reize des Riechens, Schmeckens und Tastens verarbeiten, Temperatur, Druck und räumliche Orientierung wahrnehmen -, bis hin zu komplexen, für die Entwicklung und Homöostase (= Aufrechterhalten eines ausgeglichenen, relativ konstanten Zustand) unseres Organismus essentiellen regulatorischen Netzwerken.
G-Protein gekoppelte Rezeptoren („GPCR“s) – die größte und vielseitigste Familie an Membranrezeptoren
GPCRs sind eine bereits in frühen Lebensformen vorhandene Familie von Membran-Proteinen, deren räumliche Struktur ebenso wie ihr Mechanismus des Signaltransfers über die Evolution konserviert geblieben sind. GPCRs sind offensichtliche Erfolgsmodelle. Seit ihrem ersten Auftreten hat die Natur daraus tausende Isoformen für das „Empfangen und Verarbeiten“ unterschiedlichster Signale entwickelt und damit eine der größten und vielseitigsten Protein-Superfamilien geschaffen. In den meisten Tierspezies stellen GPCRs die größte Proteinfamilie dar: Im menschlichen Genom gibt es beispielsweise mehr als 800 verschiedene Gene – das sind nahezu 4 % des Genoms -, die für unterschiedliche GPCRs kodieren.
GPCRs leiten Informationen von außerhalb ins Innere der Zelle weiter. Diese Informationen werden durch ein sehr weites Spektrum an Signalen ausgelöst: durch körpereigene Signale wie z.B. Kationen, Hormone, Lipide, Zucker, Neurotransmitter, Wachstumsfaktoren, ebenso wie durch sensorische Signale aus der Umwelt wie z.B. Photonen (Licht), Geruchs- und Geschmacksstoffe. Daraus resultiert eine Schlüsselrolle der GPCRs im Großteil aller unserer physiologischen Funktionen und ebenso in der Wahrnehmung unserer Umwelt.
Folgerichtig sind inadäquate Expression und/oder Dysfunktion des einen oder anderen Rezeptors auch mit einer Vielzahl an Krankheiten assoziiert. Es verwundert nicht, daß GPCRs zu den erfolgreichsten Zielstrukturen für wirksame Therapeutika wurden: Bis zu 50 % aller heute verschriebenen Arzneimittel wirken über GPCRs, darunter fallen so bekannte Klassen wie z.B. Beta-Blocker, Antihistaminika, Antipsychotika, zahlreiche Schmerzmittel u.v.a.m.
Als Sensoren für die Umwelt kommt unseren Geruchsrezeptoren offensichtlich besondere Bedeutung zu: Nahezu 400 GPCRs – d.i. die Hälfte aller humanen GPCRs – sind Rezeptoren für Geruchsstoffe, 28 GPCRs vermitteln Geschmacksempfindungen, 4 Rezeptoren optische Signale.
Wie funktionieren GPCRs?
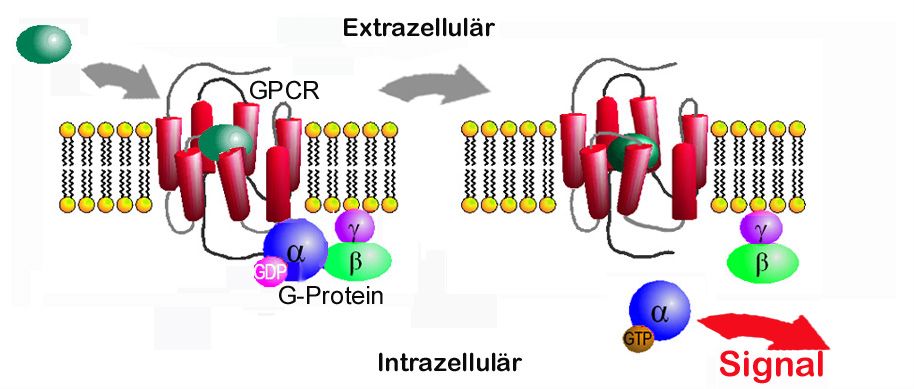
GPCR’s sind in der Zellmembran eingebettet, durchdringen diese von der Außenseite bis ins Zellinnere und leiten Signale von außerhalb der Zelle ins Zellinnere weiter. Vereinfacht dargestellt: Jeder GPCR besitzt in seinem zur Außenseite orientierten Teil eine für ein bestimmtes Molekül (Ligand) hochspezifische „Bindungstasche“. Wenn ein Ligand in diese Tasche genau hineinpaßt, bewirkt dies eine Konformationsänderung des Rezeptors, auch in seinem innerhalb der Zelle befindlichen Teil. An dieses intrazelluläre Ende koppelt ein sogenanntes G-Protein, welches durch die Strukturänderung des Rezeptors aktiviert wird, seinerseits nun Effektorproteine aktiviert und damit eine spezifische, vielfach verstärkte Kaskade von Signalen auslöst, die schließlich zur zellulären Antwort führen (Abbildung 1).
Abbildung 1: Struktur eines GPCR und Signaltransfer
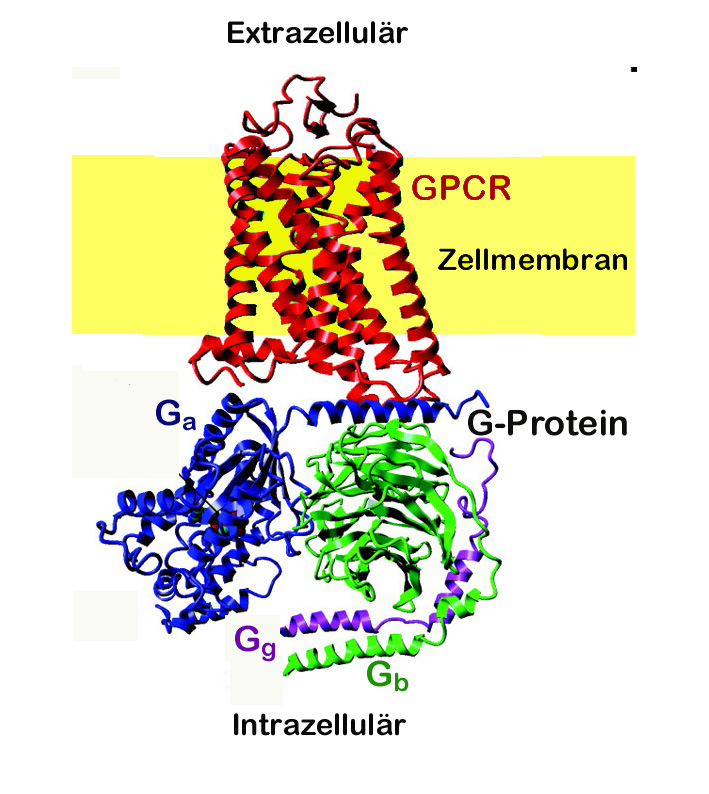 |
Links: Kristallstruktur des aktivierten beta-2 -Adrenozeptors, an den ein G-Protein (Gs) bindet. Der Rezeptor durchquert mit helikalen Segmenten 7x die Zellmembran. Der Ligand bindet im oberen Teil der Transmembran-domänen. An das intrazelluläre Ende ist das aus 3 Untereinheiten (blau, grün, lila) bestehende Gs-Protein assoziiert. (Quelle: PDB 3SN6) Unten: Bindung eines Liganden bewirkt Struktur-änderung des Rezeptors, die eine Aktivierung des G-Proteins auslöst (Austausch GDP gegen GTP) und dessen Dissoziierung in Untereinheiten, die ihrerseits unterschiedliche Signalkaskaden auslösen. (modifiziert nach Sven Jähnichen GNU-FDL, 2006) |
 |
|
Sensoren nach dem Vorbild der Natur
Weltweite Grundlagenforschung an einer Vielzahl an GPCRs hat deren essentielle Rolle in physiologischen Funktionen nachgewiesen und die wichtigsten Schritte in den von GPCRs ausgelösten Signalkaskaden auf molekularer Ebene aufgeklärt. Die Ergebnisse dieser Forschungen haben u.a. zu zahlreichen neuen, hochspezifischen und hochwirksamen Arzneimitteln geführt (siehe oben) und zu innovativen therapeutischen Konzepten.
Aufbauend auf den Erkenntnissen zur Funktionsweise von GPCRs lassen sich diese Prinzipien biomimetisch – d.i. die Natur nachahmend – auch in technische Anwendungen umsetzen. Besondere Bedeutung kommt hier Sensoren zu, die nach dem Vorbild der Photosynthese Lichtenergie in elektrische Energie umwandeln können – etwa die „elektrochemische Farbstoff-Solarzelle“ von Michael Graetzel (http://www.heise.de/tr/artikel/31-Prozent-Wirkungsgrad-sind-mit-intensiver-Forschung-drin-1027456.html) – oder nach dem Vorbild des Riechvorgangs als „künstliche Nase“ unterschiedlichste Gerüche erkennen. Die Anwendungsmöglichkeiten derartiger künstlicher Nasen sind enorm: von der Qualitätskontrolle unterschiedlichster Produkte, Erkennung von Schadstoffen in der Umwelt, Diagnostik von Krankheiten bis hin zu kriminaltechnischen Anwendungen (Aufspüren von Drogen, explosiven Stoffen).
Zu diesem neuen und innovativen Gebiet biomimetischer Anwendungen erscheint, wie schon eingangs angekündigt, ein detaillierter Bericht über die Grundlagen zur Schaffung künstlicher Nasen und den Status der technischen Umsetzung von Wolfgang Knoll (wissenschaftlicher Geschäftsführer des Austrian Institutes of Technology (AIT)) ab nächster Woche.
Glossar
- GPCR
- G-Protein gekoppelter Rezeptor
- G-protein
- Guanin-Nukleotid (G) bindendes Protein. Ein aus drei unterschiedlichen Untereinheiten bestehendes Protein, welches in seinem „Ruhezustand“ Guanosindiphosphat (GDP) bindet, in der aktivierten Form GDP durch Guanosintriphosphat (GTP) austauscht. Dieser Austausch führt zur Dissoziation der α-GTP-Untereinheit vom Rest des G-Proteins, die dann auf (membrangebundene) Effektoren stößt – Proteine und diese anschaltet.
- GTP
- Guanosintriphosphat, energiereiches Molekül, das aus der Purinbase Guanin, dem Zucker Ribose und drei Molekülen Phosphat zusammengesetzt ist. Hydrolyse eines Phosphatrestes zu GDP setzt Energie frei. GTP wird im Citrat-Cyclus erzeugt und findet als Energieüberträger Anwendung u.a. im Signaltransfer via G-Proteine, in der Protein Biosynthese, in der Polymerisierung von Tubuli, etc.
- Printer-friendly version
- Log in to post comments