Do, 04.01.2018 - 07:58 — Peter Schuster 
![]()
Von der Vermehrung von Populationen in einer Welt mit endlichen Ressourcen zu Darwins Prinzip der natürlichen Auslese und über die Vereinigung dieses Selektionsprinzips mit der Mendelschen Genetik zu einer biologischen Evolutionstheorie spannt sich ein weiter Bogen bis hin zu den heutigen Vorstellungen über die Mechanismen, die den Evolutionsprozessen zugrundeliegen. Der theoretische Chemiker Peter Schuster beschäftigt sich seit mehr als vierzig Jahren mit fundamentalen Fragen zu diesen Mechanismen und hat wesentlich zum Modell des "Hyperzyklus" und der "Quasispezies" beigetragen.*
Vermehrung in einer endlichen Welt
Spätestens seit Fibonaccis Hasenmodell (um ca. 1227) ist allen Naturalisten, Ökonomen und Philosophen geläufig, dass bei unbegrenzten Nahrungsvorräten die Populationsgrößen von Tieren von Generation zu Generation wie geometrische Reihen zunehmen (Abbildung 1). Der englische Nationalökonom Thomas Robert Malthus hat im Jahre 1798 diese Überlegungen auf die menschliche Population und die ökonomischen Konsequenzen unbeschränkten Bevölkerungswachstums angewendet. Gemäß einer geometrischen Reihe (einem ihr entsprechenden exponentiellen Wachstum), konsumiert eine derart wachsende Population alle Ressourcen eines endlichen Ökosystems bis Hungerkatastrophen drohen. Es gibt auch andere Auswirkungen einer Überbevölkerung - ein aktuelles Beispiel ist der anthropogene Anteil am Klimawandel.
Die von Malthus angesprochenen, durch Verknappung von Ressourcen entstehenden Probleme haben Wissenschaftler - darunter Charles Darwin und der belgische Mathematiker Pierre-François Verhulst - entscheidend beeinflusst. Verhulst kam auf die Idee, in die Gleichung für exponentielles Wachstum eine Beschränkung in Form einer endlichen Tragfähigkeit (C: Capacity) des Ökosystems einzuführen (Abbildung 1).
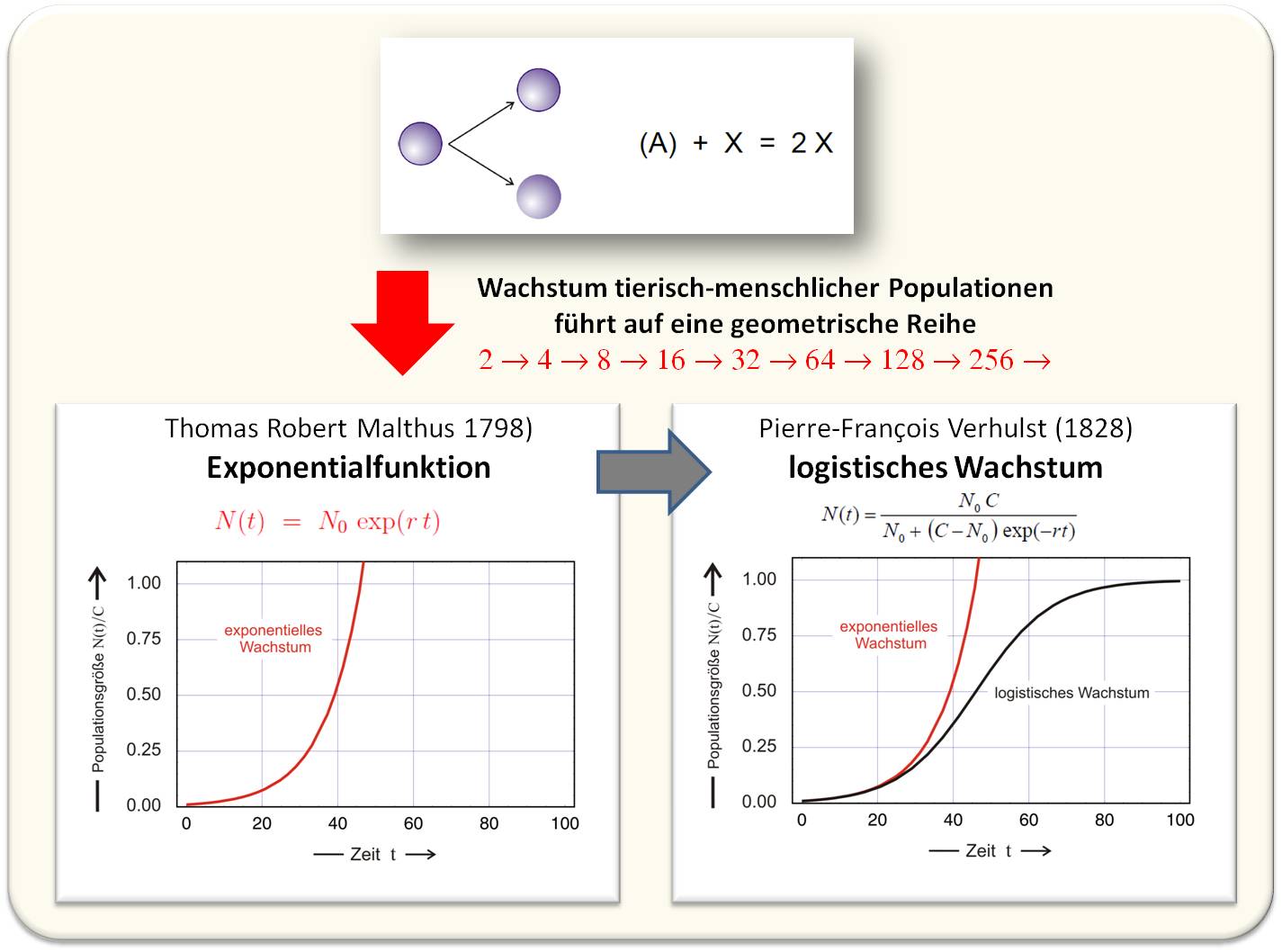 Abbildung 1. Vermehrung in einer Welt mit endlichen Ressourcen. (N = Zahl der Individuen, die im betrachteten Ökosystem leben.)
Abbildung 1. Vermehrung in einer Welt mit endlichen Ressourcen. (N = Zahl der Individuen, die im betrachteten Ökosystem leben.)
Darwin und das Selektionsprinzip
Vom logistischen Wachstum zu einer Selektion, die zur natürlichen Auslese führt, ist nur ein kleiner Schritt: wie bei Verhulst besteht die Population zwar aus einer einzigen Spezies, aber diese ist nicht homogen, sondern in Subspezies/Varianten aufgespalten, die sich in ihren Fitnesswerten (s.u.) unterscheiden. Davon bleibt die Kapazität eines Ökosystems unberührt- d.i. alle Subspezies zusammen können im Maximum nicht mehr Individuen umfassen als eine einzige. Aber es kommt zur Konkurrenz zwischen den einzelnen Subspezies und jene Subspezies mit im Mittel den meisten Nachkommen wird schließlich als Einzige überbleiben - die natürliche Auslese (Englisch: „Natural selection“ oder "Survival of the fittest"). Fitness bezieht sich hier ausschließlich auf die Zahl der fruchtbaren Nachkommen in den Folgegenerationen und hat nichts mit allgemeinem Erfolg im Leben, körperlicher Tüchtigkeit oder Durchsetzungsvermögen zu tun. Während des Selektionsprozesses nimmt die mittlere Fitness der Population laufend zu, präzise ausgedrückt niemals ab, wie durch elementare Mathematik bewiesen werden kann.
Einem mathematischen Modell wäre Darwin (1809 - 1882) sicher skeptisch bis ablehnend gegenübergestanden.
Was war aber dann seine geniale Leistung?
Versetzen wir uns dazu in die Welt eines Naturalisten des 19. Jahrhunderts, der für seine Beobachtungen bloß seine Augen und das Lichtmikroskop zur Verfügung hatte. Was dieser sah, war ein unwahrscheinlicher Reichtum an verschiedenen Formen und Funktionen der Lebewesen - Mikroben, Pilze, Pflanzen und Tiere. Für Darwin war seine Weltreise auf der HMS Beagle entscheidend, die ihn unter anderem zu den Galapagosinseln führte, wo er „Evolution in action“ beobachten konnte. Auch Alfred Russel Wallace, ein Zeitgenosse Darwins, muss hier erwähnt werden, der - basierend auf beobachteten Anpassungen von Tierarten im Amazonasgebiet und im Malaiischen Archipel und völlig unabhängig von Darwin - eine äquivalente Theorie der natürlichen Auslese entwickelt hat. Darwin und Wallace sammelten In akribischer Art und Weise Material über nahe verwandte biologische Arten und kamen zu dem Schluss, dass diese ihr heutiges Aussehen durch denselben, auf drei Säulen beruhenden Mechanismus erhalten hatten, durch:
- Vermehrung und Vererbung – die Kinder ähneln ihren Eltern,
- Variation – die Kinder sehen nicht genauso wie ihre Eltern aus – und
- Beschränktheit aller Ressourcen.
Die Bedingungen (i) und (iii) führen zwanglos zum Prinzip der natürlichen Auslese. Über Vererbung, die Mechanismen der Variation von Erscheinungsbild und Eigenschaften von Organismen, existierten damals aber nur hochspekulative Vorstellungen.
Für eine natürliche Auslese- im Sinne einer Optimierung der mittleren Fitness einer Population - müssen einige Voraussetzungen erfüllt sein. Im Allgemeinen dauert der Selektionsprozess in kleinen Populationen weniger lang, weshalb diese von Vorteil sind. Andrerseits benötigt man auch hinreichend große Subpopulationen: Ist die Subpopulation, zu der die Variante mit größter Fitness gehört, sehr klein, spielen mehr oder minder zufällige Prozesse oder unkontrollierte Schwankungen eine wichtige Rolle. Allerdings ist es für den Evolutionsprozess als Ganzes bedeutungslos, ob sich die beste, die zweitbeste oder die drittbeste, etc., Variante durchsetzt, solange echte Verbesserungen eintreten.
Mendel und die Variation durch Vererbung
Ein Grundpfeiler der Evolution wurde bisher noch nicht behandelt: Variation durch Vererbung. Darwins Vorstellungen von Vererbung waren schlichtweg falsch; entweder kannte er Gregor Mendels Arbeiten nicht oder hielt sie irrelevant für die Vorstellungen der biologischen Evolution.
Mendel (1822 - 1884) konnte durch die Interpretation sorgfältiger Versuche und die Anwendung von Mathematik, insbesondere von Statistik, seine Regeln für die Vererbung herleiten (Abbildung 2). Vererbung erfolgt in einzelnen Merkmalen und für jedes dieser Merkmale besitzt jedes Individuum zwei Träger. Sind die Träger gleich, spricht man von Reinerbigkeit andernfalls ist das Individuum mischerbig:
- Uniformitätsregel: In der ersten Generation (F1) sind alle Nachkommen von zwei verschieden, reinerbigen Elternteilen (P) gleich und mischerbig.
- Segregationsregel: Werden zwei Individuen der ersten Generation miteinander gekreuzt, so treten in der zweiten Generation (F2) alle Kombinationen auf und zwar je ein Enkel mit den beiden reinerbigen Formen (P) sowie die beiden mischerbigen Formen (F1)
- Unabhängigkeitsregel: Zwei oder mehrere Merkmale werden unabhängig voneinander vererbt.
Wie sich bald herausstellte, hat die Regel (iii) nur eingeschränkte Gültigkeit und ist nur dann erfüllt, wenn die Träger auf dem Genom sehr weit voneinander entfernt situiert sind.
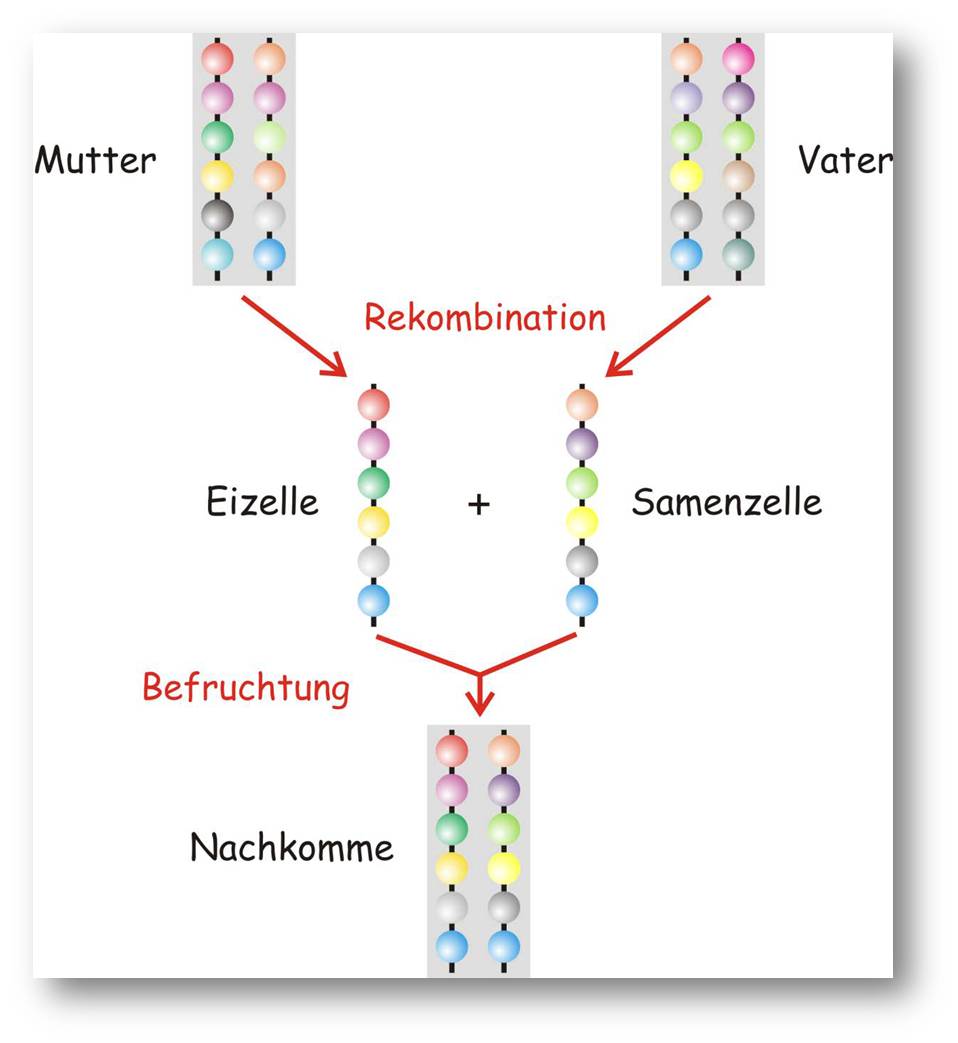 Abbildung 2. Rekombination und Mendels Vererbungsgesetze. Bei der Bildung von Ei- und Samenzellen durch Reduktionsteilung (Meiose) wird das diploide Erbgut, in welchem jedes Gen in zwei Exemplaren enthalten ist, in je zwei haploide Genome aufgeteilt, wobei die Auswahl, welche der beiden Genkopien der Mutter oder des Vaters in die das Genom der haploide Zelle aufgenommen wird, zufällig erfolgt. Bei der Befruchtung der Eizelle durch eine Samenzelle werden die beiden haploiden Genome zu einem diploiden Genom zusammengeführt.
Abbildung 2. Rekombination und Mendels Vererbungsgesetze. Bei der Bildung von Ei- und Samenzellen durch Reduktionsteilung (Meiose) wird das diploide Erbgut, in welchem jedes Gen in zwei Exemplaren enthalten ist, in je zwei haploide Genome aufgeteilt, wobei die Auswahl, welche der beiden Genkopien der Mutter oder des Vaters in die das Genom der haploide Zelle aufgenommen wird, zufällig erfolgt. Bei der Befruchtung der Eizelle durch eine Samenzelle werden die beiden haploiden Genome zu einem diploiden Genom zusammengeführt.
Mendel hat viele Tausende von Einzelbefruchtungen durchgeführt, um seine Regeln abzuleiten - alle zahlenmäßigen Aussagen gelten im Mittel großer Zahlen, haben daher mit statistischer Analyse zu tun, die für die Naturwissenschaft in der Mitte des 19. Jahrhunderts durchaus noch ungewöhnlich war.
Obwohl die Menschen seit dem Beginn ihrer Sesshaftigkeit in der Jungsteinzeit – etwa vor 12000 Jahren – begannen, Tiere und Pflanzen für ihre Zwecke zu verändern, gibt es systematische Verfahren erst seit der zweiten Hälfte des 19. Jahrhunderts. Ohne die Kenntnis genetischer Aspekte wurden Tiere nur nach äußeren Merkmalen für die Paarung ausgewählt und die Erzeugung verbesserter oder neuer Pflanzensorten erfolgte durch künstliche Selektion gewünschter Formen und blinde Kreuzung mit anderen Sorten. Erst an der Schwelle zum 20. Jahrhundert wurde die Tragweite von Mendels Arbeiten als Grundlage der Vererbung erkannt.
Der dänische Botaniker Wilhelm Johannsen prägte 1909 den Begriff des Gens, in welchem er allerdings eine abstrakte Vererbungseinheit ohne jegliche physische Realität sah. In der Folge entstand die Genetik als ein eigener Wissenschaftszweig der Biologie und ihre Anwendung stellte die Pflanzen- und Tierzüchtung auf eine wissenschaftliche Basis. Das abstrakte Bild wurde durch die Molekularbiologie korrigiert als es gelang den Träger der genetischen Merkmale in Form eines Desoxyribonukleinsäuremoleküls (DNA) mit einer wohl definierten physikalischen Struktur zu identifizieren.
Die synthetische Evolutionstheorie
Evolutionstheorie und Genetik standen lange Zeit im Clinch und es waren die Populationsgenetiker, Ronald Fisher, J.B.S. Haldane und Sewall Wright, denen um etwa 1930 die Synthese von Mendelscher Genetik und Darwinscher natürlicher Selektion in Form einer mathematischen Theorie gelang. In der Biologie beendete die sogenannte synthetische Evolutionstheorie erst mehr als zehn Jahre später den Streit. Berühmte Vertreter waren Theodosius Dobzhansky und Ernst Mayr.
Trotz unleugbarer Erfolge dieser synthetischen Theorie blieben grundlegende Probleme offen. Vor allem fehlte ein zufriedenstellender Mechanismus für die Entstehung echter Neuerungen durch den Evolutionsprozess. Rekombination (Abbildung 2) kann zwar eine gewaltige Vielzahl von Varianten bestehender Organismen erzeugen aber keine echten Innovationen. Auch evolvieren Organismen, die sich asexuell - ohne obligate Rekombination - vermehren, ebenso perfekt wie sexuell reproduzierende höhere Lebewesen. Die Mutation – in der ersten Hälfte des 20. Jahrhunderts noch vollkommen unverstanden hinsichtlich des Mechanismus ihrer Entstehung – konnte zwar für kleine Innovationsschritte und eine Optimierung von Eigenschaften verantwortlich gemacht werden, die Artenbildung erschien den Biologen aber stets als großer Sprung in den Eigenschaften der Organismen. Die meisten Evolutionsbiologen lehnten große Sprünge ab, da sie an die kreationistisch geprägten "Saltationstheorien" des 19. Jahrhunderts vor Darwin erinnerten.
Die Diskussion über die Geschwindigkeit der Evolution – langsam und graduell in kleinen Schritten oder sprunghaft, plötzlich und in großen Schritten – ging weiter. Geblieben von dieser Debatte ist die Einsicht, dass Evolution auf der makroskopischen und direkt beobachtbaren Ebene mit sehr unterschiedlichen Geschwindigkeiten stattfinden kann. In Evolutionsexperimenten mit Bakterien konnten solche Ungleichmäßigkeiten in den Prozessgeschwindigkeiten unmittelbar beobachtet werden. Computersimulationen der Evolution von RNA-Molekülen zeigten ebensolche Sprünge, die in diesem besonders einfachen Fall auch molekular interpretiert werden konnten.
Die Brücke von der Chemie zur Biologie
Bereits im 19. Jahrhundert begannen Chemiker biologische Prozesse mit physikalisch-chemischen Methoden zu studieren; Chemie und Biologie begannen als Biochemie miteinander zu verschmelzen. Anfangs galt das Interesse der Biochemiker den „Fermenten“ (Enzymen), hochspezifischen und überaus effizienten biologischen Katalysatoren, Proteinmolekülen, deren Wirkungsweise wir heute auf der Ebene ihrer molekularen Strukturen verstehen.
Als Meilenstein im Verstehen der evolutionsbiologischen Prozesse wird zurecht der auf Röntgenstrukturanalyse aufbauende Vorschlag einer molekularen Struktur für das DNA-Molekül (in der B-Konformation) durch James D. Watson und Francis H.C. Crick angesehen. Die doppelhelikale Struktur mit den nach innen gerichteten Nukleotiden, die sich eindeutig zu komplementären Basenpaaren zusammenfinden (Abbildung 3), klärte mehrere offene Fragen der Evolutionsbiologie mit einem Schlag:
- DNA-Moleküle sind Kettenpolymere wie auch viele andere Polymere, beispielsweise die Proteine. Das besondere an der DNA-Struktur ist eine Geometrie, die es gestattet die Reihenfolge der Substituenten an der Kette {A,T,G,C} abzulesen, wodurch das Molekül zur Kodierung von Nachrichten in der Art eines Informationsträgers geeignet ist.
- Die Paarungslogik, A=T und G≡C, verbindet jede lineare Folge von Buchstaben mit einer eindeutig definierten Komplementärsequenz; und man kann daher von einer geeigneten Struktur zur Kodierung von im Nukleotidalphabet {A,T,G,C} digitalisierten Nachrichten sprechen.
- Das DNA-Molekül besteht aus zwei Strängen; jeder für sich enthält die volle Information für das zweisträngige Gesamtmolekül und kann unzweideutig zu einem kompletten DNA-Molekül ergänzt werden. Dieser Sachverhalt suggeriert unmittelbar einen Kopiermechanismus (wie dies Watson und Crick in ihrer berühmten Publikation in Nature auch erkannten)
- Die DNA-Doppelhelix lässt ebenso unmittelbar eine möglichen Mechanismus für Mutationen erkennen, der in dem Fehleinbau eines einzigen Nukleotids besteht und der sich später auch tatsächlich in Form der Punktmutation als einfachste Veränderung der Nukleotidsequenz herausgestellt hat.
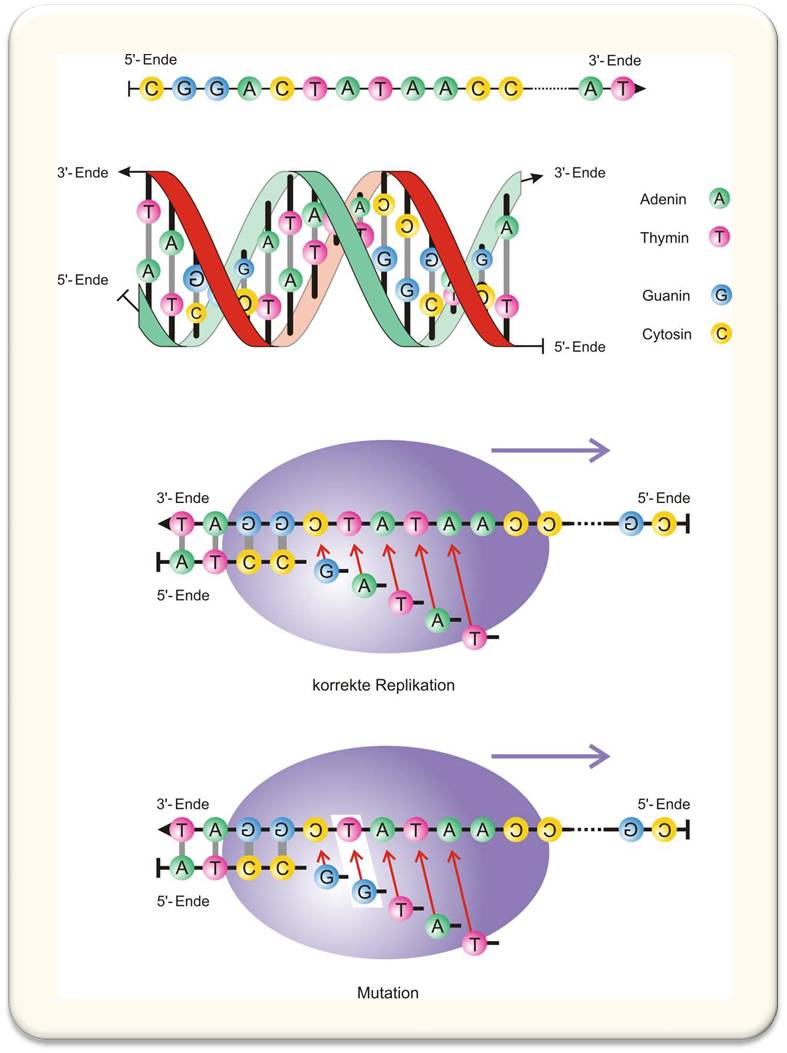 Abbildung 3. Struktur der Desoxyribonukleinsäure (DNA) und Kopieren von Molekülen. Die DNA ist ein unverzweigtes Kettenmolekül, an das vier verschiedene Nukleotidbasen A, T, G und C, angehängt sind. Die DNA-Doppelhelix (zweites Bild von oben) besteht aus zwei in verschiedene Richtungen laufenden Einzelsträngen mit den Seitenketten im Inneren der Helix. Die beiden Stränge sind über ihre Seitenketten durch spezifische zwischenmolekulare Bindungen aneinander geknüpft, wobei nur zwei komplementäre Paarungen, A=T und G≡C, auftreten. Die Komplementarität der Nukleotidbasenpaare gestattet es, einen Einzelstrang eindeutig zu einem Doppelstrang zu ergänzen; damit ist ein Weg zur Vervielfältigung von DNA-Molekülen vorgezeichnet (zweites Bild von unten). Mutationen kommen beispielsweise durch den zeitweise vorkommenden Fehleinbau von Nukleotidbasen zustande (unterstes Bild).
Abbildung 3. Struktur der Desoxyribonukleinsäure (DNA) und Kopieren von Molekülen. Die DNA ist ein unverzweigtes Kettenmolekül, an das vier verschiedene Nukleotidbasen A, T, G und C, angehängt sind. Die DNA-Doppelhelix (zweites Bild von oben) besteht aus zwei in verschiedene Richtungen laufenden Einzelsträngen mit den Seitenketten im Inneren der Helix. Die beiden Stränge sind über ihre Seitenketten durch spezifische zwischenmolekulare Bindungen aneinander geknüpft, wobei nur zwei komplementäre Paarungen, A=T und G≡C, auftreten. Die Komplementarität der Nukleotidbasenpaare gestattet es, einen Einzelstrang eindeutig zu einem Doppelstrang zu ergänzen; damit ist ein Weg zur Vervielfältigung von DNA-Molekülen vorgezeichnet (zweites Bild von unten). Mutationen kommen beispielsweise durch den zeitweise vorkommenden Fehleinbau von Nukleotidbasen zustande (unterstes Bild).
Weitere grundlegende Entdeckungen betrafen die Biochemie der Übersetzung der genetischen Information von Nukleinsäuren in Proteine, die vorerst als die einzigen wesentlichen Funktionsträger in den Zellen angesehen wurden. Gene waren nun keine abstrakten Einheiten mehr sondern konnten mit Sequenzabschnitten auf der DNA identifiziert werden. Die Entwicklung effizienter und preisgünstiger Verfahren der DNA-Sequenzanalyse ermöglicht es, vollständige DNA-Sequenzen einzelner Gene und ganzer Organismen zu bestimmen und zu vergleichen und eröffnet ungeahnte Möglichkeiten von der Biologie und Medizin bis zur Forensik.
Für die Evolutionstheorie besonders bedeutsam war die (an Hand von Aminosäuresequenzen in Proteinen) erfolgte Entdeckung der neutralen Evolution durch den Japaner Motoo Kimura. Mit Hilfe eines theoretischen Modells sowie Sequenz- und Funktionsvergleichen von Proteinen konnte er zeigen, dass Selektion auch in Abwesenheit von Fitnessdifferenzen eintritt. Selektion ist dann das Ergebnis eines stochastischen Prozesses: Welche Variante selektiert wird, kann nicht vorhergesagt werden und wir haben es dann nicht mit „Survival of the fittest“, den es ja nicht gibt, sondern mit der Tautologie „Survival of the survivor“ zu tun.
Evolutionsexperimente
Wie natürliche Auslese erfolgt, kann durch einfache Experimente mit RNA-Viren oder RNA-Molekülen untersucht werden. Die ersten einfach interpretierbaren Studien gehen auf den US-amerikanischen Biochemiker Sol Spiegelman zurück. Gleichzeitig entwickelte Manfred Eigen eine molekulare Theorie der Kinetik von Evolutionsvorgängen - zwei Ergebnisse hatten weitreichenden Einfluss auf das Verstehen der Evolution:
- Stationäre Populationen bestehen - insbesondere bei hinreichend hohen Mutationsraten - nicht nur aus einem einzigen Genotyp sondern aus einer Familie von nahe verwandten Genotypen, die Quasispezies genannt werden und aus der selektierten Sequenz sowie ihren häufigsten Mutanten bestehen, und
- In den meisten Fällen gibt es eine Fehlerschranke, welche darin zum Ausdruck kommt, dass Systeme mit Mutationsraten über einem kritischen Wert keine stabilen Zustände ausbilden können, sondern in der Art eines Diffusionsprozesses durch den Sequenzraum wandern. Die Fehlerschranke liegt etwa beim reziproken Wert der Genomlänge und dies ergibt bei Viren eine Fehlerrate von 1:10 000 und beim menschlichen Genom einen Wert von 1: 3 Milliarden.
Quasispezies bilden sich u.a auch bei Virus-Infektionen aus; die so entstehenden Viruspopulationen sind spezifisch für das Virus und für den infizierten Wirt. Dementsprechend wurde die Quasispeziestheorie auch zur Entwicklung von neuen Strategien gegen Virusinfektionen eingesetzt. Der Grundgedanke ist, durch eine Erhöhung der Mutationsrate mittels Gaben geeigneter Pharmaka die Viruspopulation zum Aussterben zu bringen und dies entweder durch Erhöhen des Anteils an letalen Varianten oder durch Überschreiten der Fehlerschranke.
Selektion in vitro wird heute auch zur „Züchtung“ von Molekülen mit vorgegebenen Eigenschaften angewandt: erfolgreiche Beispiele sind Proteine und RNA- oder DNA Moleküle.
Molekulare Genetik des 21. Jahrhunderts
Fast bis zum Beginn des 21. Jahrhunderts wurde das molekularbiologische Wissen um die Genetik von der Viren- und Bakteriengenetik bestimmt. Dass diese für höhere Organismen nicht zutrifft, diese keine "Riesenbakterien" sind, wird inzwischen klar gezeigt: es gibt grundlegende Unterschiede in den Regulationsmechanismen der Genexpression, und die RNA spielt darin eine fundamentale Rolle. Nach Meinung des Australischen Biologen John Mattick eignen sich die gut erforschten bakteriellen Mechanismen der Genregulation eben nur für kleine Gennetzwerke von bis zu einigen Tausend Genen - d.i. etwa die Länge der Bakteriengenome. Noch ungeklärt ist, ob der Großteil der nicht-translatierten DNA auch funktionslos - "junk-DNA" -ist, oder ob alle Genomabschnitte für die Regulation der komplexen Funktionen des Vielzellerorganismus gebraucht werden.
Was ist Epigenetik?
Genauere Untersuchungen zur Vererbung von Genexpression und Genregulation in einer Vielzahl von Organismen haben große Unterschiede aufgezeigt. Viele der schwer oder gar nicht erklärbaren Phänomene, die früher als Epigenetik abgetan wurden, beginnen wir nun auf der molekularen Ebene zu verstehen. Genaktivitäten sind abhängig von „Markern“ die, ohne die DNA-Sequenzen zu verändern, angebracht und abgenommen werden können. Solche Marker können von mehr oder weniger weit zurückliegenden Vorfahren und auch durch Umwelteinflüsse angebracht worden sein, haben typischerweise eine Lebensdauer von einigen Generationen und gehen dann wieder verloren. Eine andere, häufige Form des Abschaltens von Genen bedient sich teilweise sequenzgleicher RNA-Moleküle. Eine dem heutigen Wissensstand entsprechende Definition von Epigenetik wäre: „Die Erforschung von Phänomenen und Mechanismen, die erbliche Veränderungen an den Chromosomen hervorrufen und die Aktivität von Genen beeinflussen, ohne die Sequenz der DNA zu verändern.“
Sicherlich haben die künftigen Forschungen auf dem Gebiet der Molekulargenetik noch viele Überraschungen für uns bereit.
Was von Darwin 158 Jahre nach der „Origin of Species“ geblieben ist
Darwin hat mit seinem „Baum des Lebens“, der als einzige Zeichnung in der „Origin of Species“ vorkommt, als erster klar zum Ausdruck gebracht, dass alle heutigen irdischen Lebewesen von einem einzigen Urahn, einer Urzelle abstammen. Diese Vorstellung Darwins hat die Grundlage für die Erstellung von Stammbäumen (Phylogenie) - auf der Basis von DNA-Sequenzvergleichen - gelegt, ohne die eine moderne Evolutionsbiologie nicht mehr auskommen könnte.
Hinsichtlich der Bedeutung Darwins für die heutige Evolutionsdynamik, können wir uns auf das Selektionsprinzip - die natürliche Auslese - und seine universelle Gültigkeit beschränken: die Fitness zählt ja nur Individuen und ist daher unabhängig vom komplexen inneren Aufbau der Organismen. Hinsichtlich der Vorstellungen von Variation und Vererbung hatte Darwin kein derzeit vertretbares Modell vor Augen.
Die Komplexität der Lebewesen nimmt während der biologischen Evolution nicht graduell, sondern sprunghaft in großen Übergängen, den „Major transitions“ zu. Zurzeit sind diese nur soweit verstanden sind, als man plausibel machen kann, dass neben dem Darwinschen Prinzip auch andere Mechanismen wirksam sind. Dabei finden sich kleinere Einheiten zu regulierten größeren Verbänden zusammen, und vormals selbständige Elemente verlieren zumindest teilweise ihre Unabhängigkeit. Beispiele sind Übergänge von:
- RNA-Welt zu DNA & Protein-Welt,
- Gen zum Genom,
- Einzeller zum Vielzeller,
- solitären Tieren zu Tiergesellschaften,
- Primatengesellschaften zu menschlichen Kulturen.
Zusätzlich zur Evolution durch Variation und Selektion kommt Kooperation zwischen Konkurrenten als neues Prinzip zum Tragen. Ein einfaches dynamisches Modell, der sogenannte „Hyperzyklus“ wurde vor vierzig Jahren entwickelt, um eine „Major Transition“, den Übergang von einer RNA-Welt zu einer DNA&Protein-Welt, plausibel machen zu können. Die einzelnen Elemente eines Hyperzyklus haben jeweils zwei Funktionen: sie sind i) als Vorlagen zu ihrer eigenen Kopierung aktiv und ii) in der Lage diese Kopierprozesse zu katalysieren. Für eine stabile Organisationsform werden die genetischen Informationsträger – in der Regel RNA-Moleküle – zu einer ringförmigen Funktionskette, dem Hyperzyklus, zusammengeschlossen.
In der makroskopischen Biologie treten solche multifunktionellen Systeme vor allem in Form der verschiedenartigen Symbiosen auf. Systematische Untersuchungen mit RNA-Molekülen haben gezeigt, dass es oft einfacher ist an Stelle von einfachen Zyklen kooperative Netzwerke zu bilden.
Um die in der Natur beobachteten Phänomene beschreiben zu können, muss Darwins Evolutionsmodell von Variation und Selektion durch die Einbeziehung von Kooperation zwischen Konkurrenten erweitert werden, und dies ist zumindest auf der Ebene der Theorie ohne große Probleme möglich.
Schlussbemerkung
Die biologische Evolution ist ebenso ein wissenschaftliches Faktum wie die Bewegung der Erde um die Sonne (vergessen wir der Einfachheit halber Einsteins Korrekturen). So wie das Ptolemäische Weltbild durch die Raumfahrt endgültig zur Fiktion wurde, so widerlegen Evolutionsexperimente das Leugnen von Evolutionsvorgängen. Zwei Unterschiede zwischen Physik und Biologie gibt es aber dennoch:
- In der Himmelsmechanik können wir Newtons Gesetze frei von Störungen durch den Luftwiderstand und anderen Komplikationen unmittelbar in Aktion beobachten, es gibt aber keine entsprechende „Himmelsbiologie“ , und
- die biologischen Studienobjekte sind ungleich komplizierter als die physikalischen.
* Kurzfassung des Schlussvortrags "Charles Darwin - gestern und heute", den Peter Schuster anlässlich des Ignaz-Lieben-Symposiums "Darwin in Zentraleuropa" (9. - 10. November 2017, Wien) gehalten hat. Die komplette Fassung (incl. 46 Fußnoten) ist auf der Homepage des Autors abrufbar https://www.tbi.univie.ac.at/~pks/Presentation/wien-lieben17text.pdf und soll 2018 auf der homepage der Lieben-Gesellschaft http://www.i-l-g.at erscheinen.
Details zu Inhalten von "Charles Darwin - gestern und heute"
Darwin Publications: Books (American Museum of History, Darwin Manuscripts Project). open access.
Robert Malthus: Eine Abhandlung über das Bevölkerungsgesetz oder eine Untersuchung seiner Bedeutung für die menschliche Wohlfahrt in Vergangenheit und Zukunft, nebst einer Prüfung unserer Aussichten auf eine künftige Beseitigung oder Linderung der Übel, die es verursacht (6. Auflage, aus dem Englischen übersetzt und frei abrufbar; Digitale Texte der Bibliothek des Seminars für Wirtschafts- und Sozialgeschichte, Uni Köln) http://www.digitalis.uni-koeln.de/Malthus/malthus_index.html
M. Eigen, P. Schuster (1979).The Hypercycle - A Principle of Natural Self-Organization. Springer-Verlag, Berlin 1979.
Artikel im ScienceBlog:
Peter Schuster (chronologisch gelistet):
- 04.03.2016: Die großen Übergänge in der Evolution von Organismen und Technologien. http://www.digitalis.uni-koeln.de/Malthus/malthus_index.html
- 23.05.2014: Gibt es einen Newton des Grashalms? http://scienceblog.at/newton-des-grashalms
- 29.11.2013. Recycling & Wachstum — Vom Ursprung des Lebens bis zur modernen Gesellschaft. http://scienceblog.at/recycling-wachstum-%E2%80%94-vom-ursprung-des-lebe...
- 19.07.2013: Können wir Natur und Evolution übertreffen? Teil 2: Zum Design neuer Strukturen. http://scienceblog.at/k%C3%B6nnen-wir-natur-und-evolution-%C3%BCbertreff...
- 24. 05.2013: Letale Mutagenese — Strategie im Kampf gegen Viren. http://scienceblog.at/letale-mutagenese-%E2%80%94-strategie-im-kampf-geg...
- 13.09.2012: Zentralismus und Komplexität. http://scienceblog.at/zentralismus-und-komplexit%C3%A4t
- 12.07.2012: Unzähmbare Neugier, Innovation, Entdeckung und Bastelei. http://scienceblog.at/unz%C3%A4hmbare-neugier-innovation-entdeckung-und-...
- 12.04.2012: Wie universell ist das Darwinsche Prinzip? http://scienceblog.at/wie-universell-ist-das-darwinsche-prinzip
- 16.02.2012: Zum Ursprung des Lebens — Konzepte und Diskussionen. http://scienceblog.at/zum-ursprung-des-lebens-%E2%80%94-konzepte-und-dis...
- Herbert Matis, 30.11.2017: Die Evolution der Darwinschen Evolution. http://scienceblog.at/die-evolution-der-darwinschen-evolution
- Richard Neher, 03.11.2016: Ist Evolution vorhersehbar? Zu Prognosen für die optimale Zusammensetzung von Impfstoffen. . http://scienceblog.at/ist-evolution-vorhersehbar-zu-prognosen-f%C3%BCr-d...
- Karl Sigmund, 01.03.2013: Die Evolution der Kooperation. http://scienceblog.at/die-evolution-der-kooperation
- Printer-friendly version
- Log in to post comments